
SUBPHYLUM VERTEBRATA
The term “vertebrate,” designates one of a group of animals with a spinal column. The Vertebrates are characterized by the occurrence of a longitudinal dorsal skeletal rachis transformed into vertebrae, sometimes cartilaginous, bearing certain phosphatic hard tissues, and having a cranium or equivalent. This subphylum is named for the vertebral column (a series of compact bony elements) that surrounds the nerve cord.
Vertebrates
are very numerous and about 40,000 living species have been described. These animals are easy to find and can be
seen with the naked eye or reside in places inhospitable to humans, like the
sea floor. Some have bones and others
cartilages. Some of them have jaws, lungs, legs, feathers etc.
General characteristics
Feeding and Digestion
Vertebrates
have a complete digestive system with complex accessory glands. They feed on a
variety of feeds and most of them have hinged jaws. The food of the earlier
vertebrates was collected by ciliary action but this habit has long been
abandoned and only in rare cases today (such as planktivorus fishes, some
whales etc).
Circulatory system
The animals
have a closed circulatory system i.e. a circulatory system in which blood flows
in blood vessels. The blood system shows two of the most vertebrate features
namely:
·
Presence
of a heart that has at least three chambers and thus provides a rapid
circulation
·
Presence
of haemoglobin within corpuscles serving to carry large amount of oxygen to
tissues. Efficiency of this system must have been a major factor in producing
dominance of these vertebrates.
Excretory system
The excretory
system of vertebrates consists of mesodermal funnels leading primarily from the
coelom to the exterior. It may be that
this type of kidney arose in connection with the abandoning of marine water for
fresh water. It is believed that all
vertebrates passed fresh water stage and it is significant that all except the
hagfish have fewer salts in their blood than there is in marine water. Elaborate
organs for regulation of osmotic pressure have been developed and they play a
large part this regulation.
Reproduction
Vertebrates
reproduces sexually, asexual reproduction don’t occur in these animals. Most of them are dioecious i.e. have separate
sex. Some are oviparous, others ovoviviparous and some viviparous.
Vertebrate evolutionary relationship
Vertebrates have a long history on
this earth -- more than 500 million, from the late Cambrian up to today.
The origin of these vertebrates is still under debate but the first
vertebrate is believed to have developed from sessile form such as ascidians
that had a free swimming larva. The
vertebrate body organization may have arisen as an adaptation for more active
life style. This probably occurred by a process known as neoteny in
which the length of the larval period is gradually extended until eventually
sexual maturity is reached without undergoing metamorphosis and then the animal
retains its larval form and features as a sexually reproducing adult. These
first vertebrates lacked jaws, like the living hagfish and lampreys. Jawed
vertebrates appeared 100 million years later.
Walter Garstang noted from study of
marine animals that both echinoderms and chordates are deuterostomes (in which the blastopore forms the anus), while most other
possible ancestors of Chordates are protostomes (the blastopore forms the mouth).
This inspired Garstang to suggest an alternate route of evolution: from
echinoderms to chordates.
There are many important differences
between chordates and echinoderms. Most importantly, adult echinoderms show
little likeness to chordates: echinoderms are radially
symmetric, possess calcium carbonate plates in their skin and have tube
feet. Garstang made the radical suggestion that perhaps it was echinoderm
larvae, not adults, which had given rise to chordates.
Echinoderm larvae, like chordates,
are bilaterally
symmetric.
Especially notable are their similarities to larvae of hemichordates, which are a step closer to
chordates as they share two of the five most commonly noted chordate
characteristics, namely a hollow neural tube and pharyngeal slits.
Garstang's idea has been expanded and
is supported by many lines of evidence. Perhaps most interesting and compelling
is the fact that some amphibians can stay in larval form and still
reach sexual maturity—this shows that echinoderm larvae could, theoretically,
have become sexually mature and simply stopped morphing into adults, instead
evolving into chordate ancestors. Species that show this refusal to leave the
larval stage include mud puppies and other salamanders, which either partially or
completely show neoteny (also called pedomorphism):
retention of juvenile traits or phenotypes after sexual maturity. These first
vertebrates lacked jaws, like the living hagfish and lampreys. Jawed
vertebrates appeared 100 million years later.
Garstang's hypothesis was
revolutionary for both its time and idea: it suggests that not only may single species
evolve, but that single life stages of species may evolve into separate
organisms. The hypothesis, which Garstang proposed in the early 20th century,
seemed far-fetched at the time of its conception and did not receive support
until after Garstang's death.
Adaptation of
vertebrates
·
Vertebrates have paired appendages that facilitate locomotion
o In fish the fins
facilitate balance, propulsion and directional movement.
o In other vertebrates fins
have developed into jointed limbs for movement and support on land. Jointed
limbs are well adapted for moving over surface that are not smooth or level.
·
Vertebrates have ciliated pharyngeal gill structures which are
modified for gaseous exchange.
o The gills are very
proficient in gas exchange and the respiratory efficiency exceeds that of
invertebrates.
o Oxygen is distributed
around the body by a well developed circulatory system.
·
Vertebrates have an endoskeleton which has several advantageous
features. It is composed of living tissue and thus grows with the animal rendering
moulting unnecessary.
o It permits greater body
size by providing a lighter support
structure.
o It provides an increased
surface area for muscle attachment permitting a wider range of movement.
·
Vertebrates have a highly developed and complex nervous system.
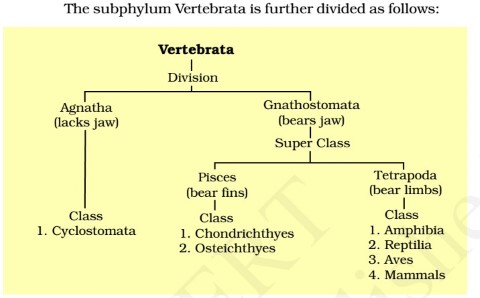
Taxonomy
Vertebrates are classified as below
├─Subphylum Vertebrata
├─Superclass Agnatha
├─Class Pteraspidomorphi
├─Class Cephalaspidomorphi
├─Superclass Gnathostomata
├─Class Placodermi — extinct (armored gnathostomes)
├─Class Chondrichthyes (cartilaginous fish)
├─Subclass Elasmobranchii — (plate gills)
├─Subclass Bradyodonti (slow tooth)
├─Class Osteichthyes (bony fish)
├─Subclass Acathodii (spiny)
├─Subclass Actinopterygii (ray-finned fish)
└─Subclass Sarcopterygii (lobe-finned fish)
├─Class Amphibia (amphibians)
├─Class Reptilia
├─Class Aves (birds)
├─Class Mammalia (mammals)
1. Superclass Agnatha
Super class Agnatha consists of an
ancient group of animals similar to fish but with some very noticeable
differences. The term Agnatha means “no jaws” in Greek. The Agnathans lack jaws
and paired fins. As with sharks (another
ancient group), the internal skeleton consists primarily of cartilage.
The only living Agnathous animals are
the Cyclostomata (ringed-mouthed) lampreys and the hagfishes but the first
vertebrates to appear in most fossil series mostly heavily armored and hence
known as ostracoderms (bonny skinned) also show Agnathous condition and have
some other features common with the cyclostomata.
General
Characteristics
· Agnathans have no jaws
· Most are bottom dwellers and wriggle
along the sea floor or streambeds.
· They don’t have any appendages,
external body parts.
·
Digestion:
Agnatha don’t have
an identifiable stomach. Their metabolism is slow and due to this they don’t
have to eat as much. They are Fluid feeders
· Osmoregulation: Some Agnathans are Osmoconformers
e.g. hagfish and some are osmoregulators. Osmoconformers normally change
the osmotic ionic conc. of their body to conform to that of the surrounding
environment thus remain isotonic.
· Reproduction: Fertilization and development is external.
There is no parental care. They are Oviparous
The first jawless fishes were Ostracoderms
(shell skin) which is an artificial designation and not a monophyletic group.
Characteristics of Ostracoderms
a) First fossils have well developed external bone, no
internal bone: Cambrian (500 mya)
b) Dominant for 100 my (gone by 380 mya)
c) First fossils: marine; later marine
and freshwater
Features of Ostracoderms
a) No jaw, muscular feeding pump
(filter feeders)
b) Body armor - true bone
c) Some had paired fin-like
appendages, but not true fins with bony support
d) Heterocercal tail
Two main Classes of Ostracoderms are Cephalaspidomorphi ( ancestor to lamprey) and Class
Pteraspidomorphi (ancestor to hagfish)
i)
Class Pteraspidomorphi
Pteraspidomorphi is an extinct class of early jawless fish which lived from the
Early Ordovician to the Late Devonian (i.e. from 470 to 370 million years ago).
The fossils show extensive shielding of the head. Some species may have lived
in fresh water. The taxon contains the subgroups Heterostraci and Thelodonti.
Pteraspidormorphs have
long been regarded as closely related -or even ancestral- to jawed vertebrates,
but the few characteristics they share with the latter are now considered as primitive
for all vertebrates.
Characteristics
· They were jawless and
possessed a massive dermal skeleton with the body divided into a scaly trunk
and a tail.
· They are characterized by
possession of an extensive dermal armor covering the head having large, median,
ventral and dorsal plates or "shields”.
· Their dermal plates and
scales are made up of acellular bone, or aspidine, but this is thought to be a
primitive condition for the dermal bone of all bony vertebrates.
· Pteraspidomorphs have no calcified endoskeleton however some show traces of
calcified cartilage.
· They have no other fin than
the caudal fin.
· The internal surface of
the dermal plates -at any rate in heterostracan- shows impressions of the
gills, brain, two vertical semicircular canal and distinctly paired nasal
capsules.
· The nasal capsules of
heterostracans seem to have opened ventrally into a separate median duct
comparable to the nasopharyngeal duct of hagfishes.
· Most Pteraspidomorphs
were marine but lived very near to the shore, in lagoons and deltas. Some
groups are thought to have been fresh water. They were certainly
bottom-dwellers, as shown by traces of abrasion of the ventral surface of their
head shield. Examples, Pteraspis, Psammosteus, etc
Limitations
of Ostracoderms
· Habitat limitations
o Restricted to benthos
o Weak swimmers due to heavy armor that
also result into inflexibility
· Food limitations
o
No
jaws - restricted to plankton, suspended organics - slow growth
2. Class
Cephalaspidomorphi
·
This
is a taxon of jawless fishes named for the Cephalaspids, a group
of Osteostracans. Most of the members of this group
are extinct; however, it interests modern biologists because it includes the lampreys.
·
Members of this taxa
have a single nostril and are jawless fish
Order Osteostraci =Cephalaspida
The order
contains fossil Agnathans that show more similarity to modern cyclostomes than
the pteraspids.
Characteristics
· First fossils have well developed
external bone, no internal bone (bone shell)
· First fossils were marine later they were
found in marine and freshwater bodies.
· No jaws, they are filter feeders
· Ventral surface of head was flat and
it was covered by small scales
· Some had paired fin-like appendages,
but not true fins with bony support
· They have a heterocercal tail
· Had paired eyes and pineal eye
· On the dorsal surface there are
sunken areas that are believed to contain electric organs.
Examples Hemicyclaspis, Kiaeraspis etc.
Order Cyclostomata
Members
include lampreys and hagfish. Lampreys belong to suborder Petromyzontidae (stone
suckers) and hagfish to suborder Myxinoidea (slime ones)
Characteristics of Cyclostomes
·
Eel-
like in structure with a size of 10-90cm in length
·
Lack
exoskeleton/scales
·
Prey
on fishes (fluid feeders).
·
Notochord persists in adults
·
Lack
jaws but have rows of horny teeth that move in circular motion
·
Axons
of neurons are unmyelinated
·
Generally
marine (some anadromous)
· Skeleton cartilaginous or fibrous
· No paired fins
· No body armor
· Single median nostril
·
Series
of round gill openings, no true gill archesl
Two Important Sub-Orders of Cyclostomata
Lampreys are
found in marine, brackish and freshwater environments. They are the most
familiar cyclostomes. Some species are parasitic, attaching their sucker-like
mouth to a fish and then using sharp teeth to rasp away at the animal's flesh.
The lamprey has a larval stage. The ammocoete larva
lives in fresh water, buried in mud. The ammocoete larva has to undergo
metamorphosis to change into an adult.
Size and Habitat
Lampreys
include marine and freshwater species example are the sea lamprey (Petromyzon
marinus), pacific lamprey (Entosphenus tridentate) and freshwater
lampreys (Entosphenus similes)
The adult
lamprey is an eel like animal about 30 cm long and lives parasitically in the
sea probably for about 2.5 years. They
then migrate to fresh water where they spawn. Fertilized eggs develop into an
ammocoete larvae which don’t have suckers and lives buried in mud feeding on
micro organisms for 5 years. After metamorphosis the young adult migrates to
the sea. This kind of migration is called anadromy and species with this kind of migration are said to be anadromous.
Body colour
Adult Lamprey
is dark on the back and white below. The body surface is smooth with no scales.
The skin is many layered. The outer most cells have striated cuticular border. The skin secretes slime with antibacterial
effects. The dermis contains pigmented cells called the chromatophores. The chromatophores
are star shaped cells whose pigment is able to migrate hence making the animal
pale or dark. This change is especially marked in larvae and it is
produced by variation in the amount of pituitary secretion
Nutrition
Lamprey larva is sedentary filter feeders but the adults are ectoparasites primarily of other
fishes. They attach to the host with the
oral disk and use the keratin plates
that cover the tongue and oral disk to rasp a hole through the skin and absorb
the fish’s blood and tissue.
Lampreys have
a simplified digestive system. Food travels from the mouth through the
oesophagus to the intestines. There is no true stomach. They have a liver, gall
bladder and bile duct but no separate pancreas.
Reproduction
Lampreys
are dioecious. They have external fertilization but there are some
modifications of the cloaca in both sex to ensure fertilization and proper
placing of eggs in a nest. The lips of the cloaca of a ripe male are united to
form a narrow penis like tube. The cloacal lips of a female are enlarged and
often red.
The generalized life cycle of lampreys is as summarized below
Parasitic forms are
anadromous- they spawn
in clear flowing streams in gravel and later juveniles migrate back to the sea.
Like salmon – they are semelparous
i.e. they invest heavily in reproduction and die after spawning
Lampreys produce small eggs (1 mm)
also one female can produce 250,000 eggs
Their larvae and juveniles are called
ammocoetes – they live in gravel and filter feed, parasitic forms undergoes
metamorphosis as they migrate downstream and adopt a parasitic lifestyle
Nonparasitic lampreys are
called brook lampreys - retain the juvenile form, filter
feed, and mature earlier and spawn fewer eggs. Brook lamprey are derived from
the parasitic form by loss of the parasitic phase. Brook
lampreys are not anadromous, they inhabit fresh water bodies.
Circulation and gaseous exchange
The blood
vascular system is arranged on the same general plan as in amphioxus but there
is a well developed heart. The heart lies behind the gills and it is three
chambered. Blood from the heart passes the ventral aorta to the gill pouches
(external gill pouches). Oxygenated blood from gill pouches is passed through
arteries to organs such as kidneys, gonads, etc. deoxygenated blood from these
organs is transported back to the heart through veins so that it may be pumped
back to the gills. Blood of lamprey contain haemoglobin as that of other
vertebrates. These animals have about 7 pairs of gills.
Excretion and Osmoregulation
The blood of
lamprey contains a much higher concentration of salts f than the surrounding
water when it is in fresh water and vice versa when in marine water. When they are
in fresh water they must deal with the tendency of water to flow in and
minerals out.
In fresh water lampreys the excess
water that moves in osmotically is removed by passing the filtrate in the
kidney and the salts are re absorbed back in the kidney tubules. Fresh water lamprey secretes a lot of urine that is hypotonic
to blood. Their bodies are also impermeable to water however not
completely impermeable; this reduces the amount of fresh water that may enter
their body. Lampreys can also take up Na+ and Cl- from
the environment by special ion uptake cells in the gills (this transport of
mineral ions is by active transport). Lampreys are therefore the osmoregulators.
Nervous system and Sense organs
Lampreys have
a well developed sensory structure including eyes and a lateral line system. In
addition to possession of eyes the lampreys have a pineal eye which is normally
associated with reproduction,
metamorphosis and change in
colouration in larvae (in day larvae becomes dark and at night pale).
The lateral line provides a distant touch sense. Mechanoreceptors detect
vibration and water movement.
The
modern hagfish are all marine, living on the bottom and usually in burrows. Rather worm-like in
appearance, the hagfish lack paired fins, only having a slight tail fin. They
have no eyes and no scales. They also have rows
of horny teeth on the tongue which rasp at the food. Surrounding the mouth is a
ring of tentacles. Hagfish produce a great deal of slime.
Habitat
and Nutrition
Hagfish lives buried in
mud or sand and eat polychaetes and other invertebrates as well as scavenging
dead and decaying fish. Have keratin
plates on teeth. They use their tongue to rasp and burrow into the body of prey
usually through the gills or anus and use their keratinized teeth to tear of
pieces of the host tissue. They don’t have a stomach
Nervous
system and sense organs
Hag fish have poor vision and the
lateral line system is greatly reduced. The eyes are functionless rudiments
though the animals are sensitive to changes of illumination through skin
receptors. They also have sensory tentacles around the mouth (barbels).
Reproduction
Hag fish are monoecious i.e. they have both male and female
sex organs but the gonads of only one sex are functional. Fertilization is external and development is direct, there is no larval stage. They produce large eggs with hooks.
Gaseous
exchange
Hagfish have up to 16 pairs of gills.
Each gill pouch connects through one or more ducts to the pharynx. The median
nostril connects to the pharynx; it allows water to pass through the pharynx
and gills while food is in the mouth. The internal gill openings combine into a
single tube connecting with the pharynx. With this structure feeding doesn’t
impede the passage of water through the pharynx and over gills.
Excretion
and Osmoregulation
Hagfish are osmoconformers; they are the only vertebrates with
body fluids isotonic to seawater. They are also the only vertebrates to have
both pronephric (a 1st functional kidney of embryo) and mesonephric
kidneys in adult. In most vertebrates the pronephric kidney degenerates in
early embryonic development and is replaced by mesonephric kidney. Mesonephros
is a full functional kidney in other fish and amphibians but it is an embryonic
kidney in birds, reptiles and mammals which is replaced by the metanephros.
An
interesting feature of hagfish
Hagfish
secrete a lot of mucus, a 50 cm hagfish can fill 8 litres bucket with mucus in
minutes. Hagfish mucous is composed of proteins and carbohydrates that bind to
water. Mucous glands are along the ventral surface
Functions of slime
·
Suffocate
prey fishes
·
Protection
from digestive enzymes
·
Discourage
other scavengers
·
Secure
burrow walls
·
Defence
from predators
Summary
of the characteristics of Lampreys & hagfish
Characteristic
|
Lamprey
|
Hagfish
|
|
Dorsal
fin
|
1 or 2
|
none
|
|
Anal fin
|
Present
|
Present
|
|
Eyes
|
well
developed
|
rudimentary
|
|
Lateral
line system
|
well
developed
|
absent
|
|
Barbels
|
Absent
|
3pairs
|
|
Nostril
location
|
on top
of head
|
on snout
|
|
External
gill openings
|
7
|
1-16
|
|
Cranium
|
Well
developed
|
Rudimentary
|
|
Osmoregulation
|
hyper- or hypoosmotic
|
isomotic
|
|
Eggs
|
small, no hooks
|
large, with hooks
|
|
Larval
stage
|
Present
|
absent
|
|
Summary of
the Key
features of Agnathans
· Jaws are absent.
· Paired fins are generally absent.
· Early species had heavy bony scales
and plates in their skin, but these are not present in living species.
· In most cases the skeleton is
cartilaginous. The embryonic notochord persists in the adult.
· Seven or more paired gill pouches are
present.
· The digestive system lacks a stomach
Superclass
Gnathostomata
Gnathostomata is the group of vertebrates with jaws. The term derives from Greek γνάθος (gnathos)
"jaw" + στόμα (stoma) "mouth".
The Gnathostomata, or gnathostomes, are the majority of the
Middle Devonian (-380 million years ago) to Recent vertebrates. They differ
from all other craniates or vertebrates in having a vertically biting device,
the jaws, which consist of an endoskeletal mandibular arch and a variety of exoskeletal
grasping, crushing, or shearing organs, i.e. the teeth, and jaw bones.
It is believed that the jaws evolved from anterior gill support arches that had acquired a new role, being modified
to pump water over the gills by opening and closing the mouth more effectively
— the buccal pump mechanism. The mouth could then grow
bigger and wider, making it possible to capture larger prey. This close and
open mechanism would with time become stronger and tougher, being transformed
into real jaws.
Characteristics
Gnathostomes are
characterized by:
· A vertically biting
device called jaws, and which is primitively made up by two endoskeletal
elements and a number of dermal elements called teeth, sometimes attached to
large dermal bones.
· The skull of a
gnathostome (here a shark), are characterized by vertically biting jaws (Y).
The gill arches (R) are situated internally to the gill filaments, and the
nasal capsules (C) open to the exterior by means of paired nostrils.
· Possess jointed, paired lateral
appendages e.g. paired pectoral fins,
limbs etc
· Gill arches lie
internally to the gills and branchial blood vessels, contrary to the gill
arches of all jawless craniates, which are external to the gills and blood
vessels.
· Possess three (3)
semicircular canals in the inner ear.
· Paired nasal sacs which are
independent from the hypophysial tube. Nasal sacs
open separately to the exterior by external nostrils. In all extant and fossil jawless
craniates, the nasal sacs, which contain the olfactory organs, open into a median
duct, the nasohypophysial duct, which takes part to the formation of the
pituitary gland and either leads postriorly to the pharynx (e.g. in hagfish) or
ends as a blind pouch (e.g. in lampreys and osteostracans). In the
gnathostomes, this pouch remains as a thin canal in the palate, the
buccohypophysial canal,
· There are numerous other
characteristics related to anatomy and physiology (e.g. myelinated nerve
fibres, sperms passing through urinary ducts, etc.), which are unique to
gnathostomes among extant craniates, but cannot be served in fossils.
Discussion of Phylogenetic Relationships
Gnathostomes have long been placed as the sister-group of the
Agnatha as Clade. Current phylogenies, however, suggest that the Agnatha are
not a clade, and that, among recent craniates, the gnathostomes are the
sister-group of the lampreys only. Among fossil craniates, the Osteostraci
share the largest number of uniquely shared derived characteristics with the
gnathostomes.
Class Placodermi
Placoderms were armored jawed fishes that first appeared about 420
million years ago (MYA) during the Silurian Period. They had diversified
dramatically by the beginning of the Devonian and came to dominate most marine
and freshwater ecosystems.
Placoderms became extinct during the early carboniferous era (355 MYA).
About 200 genera of placoderms have been discovered, with most of these
occurring during the Devonian radiations. The rapid evolution and diversity of
placoderms have made them useful in dating Devonian rocks.
Key features of Placoderms
· Placoderms (= plated skin) were named
for their heavy armor of dermal bone, which formed large shields on the head
and thorax. The rest of their
bodies was covered with small bony scales or was without dental armor.
·
They had jaws but no ability to extrude them
·
Placoderms
lacked teeth, but they had sharp bony plates that they used as teeth.
·
They
had true paired fins
·
Most
placoderms were less than 30 cm (2 feet) in length, but some members reached or
exceeded 6 m (20 ft), making them the first giants of the vertebrate lineage.
· Possess a bony internal skeleton
· Most had a heterocercal tail
Traits shared with Ostracoderms
· Negatively buoyant (due to heavy
plates)
· Occupied benthic and near-benthic
habitats (epi-benthic)
· Dorsoventrally depressed (common among
benthic fishes)
Differences from Ostracoderms
· Placoderms reached much greater sizes
o
up
to 10 m (33 feet) in length
o
Why?
- food source, mobility
· Placoderms had slightly lighter and
more flexible (articulated) armor
